~第2回:ウサギの食べ物と、恐竜の比べっこ~
こちらは前回から投稿している、2023年が卯年ということに因んだ企画、『卯年に考えるウサギみたいな恐竜 “完全版”』の続きです。
1).はじめに
前回お話したように、ウサギは哺乳類、恐竜は爬虫類とそれぞれ異なる仲間であり、これら2つの仲間は3億年以上前に分かれた遠い関係性にあるほか、それぞれの生息環境や時代も異なるため、ウサギのような体の形を持ち、ウサギのような物を食べ、ウサギのような行動をする、3つ全てが似ている“ウサギそっくり”の恐竜は発見いません。
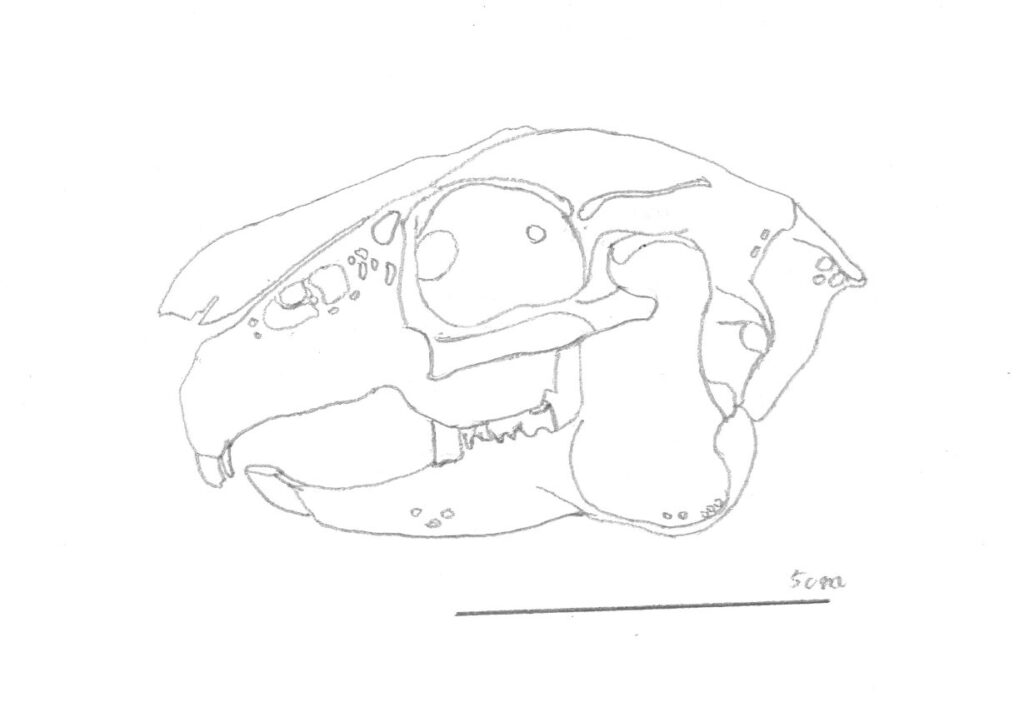
しかし、部分ごとに比べてみると似ている点もあり、第1回目の体の形の比較では、大きな前歯がウサギを思わせる、インキシボサウルスなどを紹介しました。
第2回目である今回は、「ウサギの食べ物と、恐竜との比べっこ」と題して、ウサギの食べ物と恐竜の食べ物で、似ているものがあるか見ていきます。
2).ウサギの“ウサギらしい食べ物”
ここでは前回と同様に、ウサギの特徴から整理し、今回は“ウサギらしい食べ物”について、ウサギ科全体から大雑把に見ていきます。

【ウサギの献立】
- 草木(木の葉、草、木の皮、根)
- 果実
- 種子
- まれに肉
3).どやって恐竜の食べ物を知るのか?
現生動物は行動観察*や糞分析*、胃の内容物*を調べることで食性を知ることができ、ウサギ科の動物は基本的に植物を食べており、餌が不足した場合や出産後の栄養失調*の場合など、環境や状態によっては肉を食べることがわかっています。
しかし、絶滅動物である恐竜においては、これらの手法を用いることは基本的にできません。そこで恐竜においては、歯や顎の形態から推測されており、現生動物の歯や頭部などとの形態の比較や、歯に残された摩耗痕、糞化石*などを調べることで恐竜の食性を明らかにしていますので、以降で幾つかの例を紹介していきます。

・3-1:歯から知る
比較する上で1番わかりやすい部分は歯で、これは歯が採食*に関わる器官であり、食べるものによって容易に形を変えるためです。世界で初めて科学的に研究された恐竜の1つであることで有名な、鳥脚類のイグアノドン*も、歯の形状が現生の植物食爬虫類であるイグアナ*の歯に似ていたことから、「イグアナの歯」を意味する学名*として、この名が付けられています。

イグアノドンの歯は木の葉状の形をしており、三角形の歯冠*とくびれた歯根*という基本形に、前後を縁取るようにギザギザがつきます。この形がイグアナの歯と似ているわけですが、これは咀嚼によって磨り減る前の形であり、摩耗した歯は植物食の哺乳類であるサイ*やゾウ*に似ており、実際に哺乳類でいうところの臼歯*の役割をもっていると考えられています。


・3-2:口から知る
植物食であったかは、口の形状からも推測することができ、植物食恐竜の中でも一部の基盤的なものを除いて鳥盤類の仲間たちでは、前上顎骨歯がなくなり、口先部分がざらついた面になります。また下顎の先には前歯骨*という歯がない骨が形成され、この上下の口先の歯がない部分には鳥類や、カメ類*に見られる角質*のクチバシ*があったことが知られています。

また、恐竜類で開いていた下顎の後ろ側に開いていた外下顎孔*と呼ばれる穴が小さくなるほか、顎の関節する部分が、歯列*の高さより下になっており、これらの特徴は咀嚼筋*が生む顎の閉じる力を、効率よく歯に伝えるための適応であると考えられています。この様な咀嚼に適した構造は、植物食哺乳類とは別の形で発達しており、鳥盤類の仲間が植物食恐竜の中でも、非常に特殊化していたことがわかります。
・3-3:頭から知る
植物食恐竜である竜脚類は、長い首や、全長に対して小さい頭などが特徴的です。この頭は小さいだけでなく、他の恐竜と比較しても軽いつくりをしており、このような形態を発達させたことが、首を長くさせやすかった要因のひとつであると考えられています。こういった頭は、獲物を仕留める武器としての役割を、頭に持たせている肉食恐竜では見られないものであり、口や歯など、その他の形態的な特徴と合わせることで、食性が見えてきます。

・3-4:胴か知る
植物は細胞壁*の影響で肉質のものと比べて消化しづらく、長い時間をかけて消化を行うため、植物食動物の消化管は長くなっています。また、ウシ*やキリン*などの反芻動物*では4つの胃のうち、第1胃*が内部共生生物*もつ発酵器*となっており、ここで植物を栄養として吸収できる形まで分解していますが、この様な器官を収めるために、胴は大きくなっています。

上述した竜脚類においても、現生動物と似た消化が考えられており、基本的に咀嚼はしないものの、植物を摘みとってそのまま飲み込むと、物理的な破壊を筋胃(砂嚢)か、これに似た器官でした後、長い腸で分解や発酵をしていたと考えられています。これは、胴の形から知ることができ、竜脚類の胴は太く、ここに現生の植物食動物がもつような、内部共生生物をもつ発酵器を備えた、大きな消化官が収められたと考えられています。特にティタノサウルス類*では腸骨*の前方が左右に広がり、他の竜脚類と比べても太い、樽のような胴体になっていました。

この様な樽状の幅の広い胴体は、パキケファロサウリア類*やアンキロサウリア類で見られ、同様に内部共生生物をもつ発酵器を備えた、大きな消化管が収まっていたと考えられています。鳥盤類内の別の系統であるパキケファロサウリア類やアンキロサウリア類、そして竜盤類*である竜脚類などで、似た適応が見られるのは非常に面白いです。

・3-5:腰から知る
鳥盤類の腰は竜盤類の多くと異なり、恥骨*が後方へ向いていますが、これは2足歩行を維持したまま植物を消化するのに必要な大きな内臓のスペースを確保するため起きた変化と考えられており、これによって移動能力と消化能力を両立させていたようです。因みに、鳥盤類には四足歩行の仲間も多く見られますが、この仲間が誕生した頃は全てが二足歩行でした。
・3-6:歯の摩耗から知る
この章の始めに、歯の形態から恐竜の食べ物を推測した例を紹介しましたが、歯の摩耗痕からも食性は研究されています。ティタノサウルス形類*に属する竜脚類の、歯の表面に残されていた微小摩耗痕を、様々な食性の現生トカゲ類と比較した研究では、歯の傷の深さや大きさから、竜脚類が貝殻より柔らかく、卵の殻や肉よりも硬い植物を食べていたと推定しており、食べ物の硬さが明らかにされています。

・3-7:体に残された内容物から知る
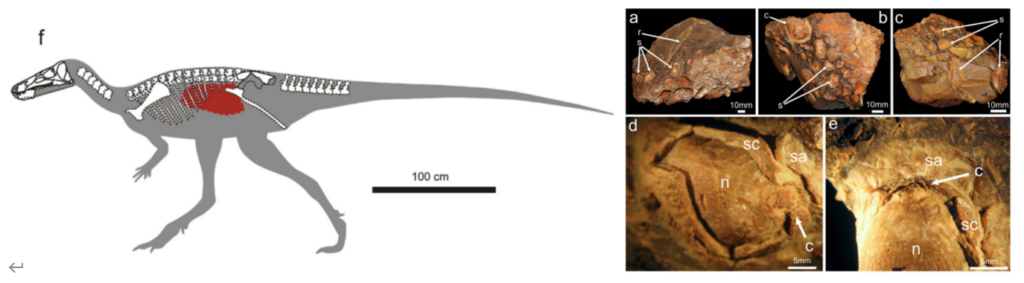
恐竜を解剖して、胃の内容物を調べることはできませんが、希に恐竜の体内に関する情報が、化石として残されている場合があり、これらの化石記録は他の記録よりも直接的に、恐竜の食性について教えてくれます。例えば、鳥脚類のハドロサウルス類では、ミイラ化した化石が幾つか発見されており、これらの化石の中には胃の内容物を残した標本があり、そこからは、小枝や実、葉などが発見されています。

ノドサウルス類の仲間であるボレアロペルタ*という恐竜でも、胃の内容物が見つかっており、そこからは1種類に由来すると考えられるシダ植物*の葉や、茎などが多く発見されたほか、裸子植物*や被子植物*、コケ*なども発見されています。また、ソテツ*などが全く見られなかったことや、木炭*が発見されており、シダ植物が火事などで環境が荒れた後に、真っ先に群生する植物であることから、食性に好みがることや、少なくとも死ぬ前の食事が、火災後に繁茂したシダ植物であった可能性が考えられています。

ただし、これらの例で発見された胃の内容物が、その恐竜が好んで食した植物だと決定し、似た形態を持つ恐竜全体も、同様の物が主な食料であったと考えるには注意が必要です。これは、同じ仲間や近い仲間の動物であっても、季節や生息地域によって食性が異なるためであり、化石として見つかった証拠が、全体に当てはまるとは断言できないためです。
例えば現生動物であるゾウの仲間でも、アフリカゾウ*は葉が多い植物食で、アジアゾウ*は草が多い植物食と、その食性は僅かながらに異なります。また、同じアフリカゾウの仲間でも季節や、生息地域によって異なり、草原地域では草を中心に、森林地域では木の芽などを中心に食しているそうです。

・3-8:うんちから知る
糞化石には多くの情報が含まれており、食していた物の破片や、バクテリア*の痕跡など、排泄した動物に関する情報や、種子や花粉など周囲の環境を知るのに役立っています。ただし、余程のことが無い限りは、その持ち主を特定するのは難しく、恐竜のものと推定されている糞化石は、大きさが当時の生態系を考えるに、恐竜以外は残し得ないものであった場合など、限られたものになります。

また、生活圏が広い動物の糞化石だった場合には、同じ地層から発見されない可能性があるほか、生活圏から離れた場所で排泄をする現生動物もいるため、この様な動物の糞化石の場合には、本来の生息地ではない可能性もあるなど、化石を解釈する際には注意が必要です。
4).植物はたくさん、植物食もたくさん
ウサギの仲間は(2)で紹介したように、基本的には植物食*であり、この点に注目するのであれば、該当する恐竜は数多く存在し、竜盤類ではフクイティタン*など長い首が特徴的な竜脚類のほかに、フクイラプトル*など肉食が多いことで有名な獣脚類にもテリジノサウルス類や、オルニトミモサウルス類*など一部の仲間が植物食であったと考えられています。

そして、フクイサウル*などの鳥脚類を含む様々な形態のある鳥盤類では、ほぼ全ての仲間が植物食であった考えられており、恐竜類の全ての系統で植物食を見ることができ、けして珍しくない共通点であると、言えるのではないでしょうか。

そもそも動物は植物食の方が多く、例えば哺乳類では、人間による影響があり、真に野生の状態を反映しているとは言えないなどの問題点はありますが、アフリカの自然動物公園における肉食動物(捕食者*)の植物食動物(被捕食者*)に対する比率は約1%であり、自然界において数が多いのは、植物食動物であることがわかります。
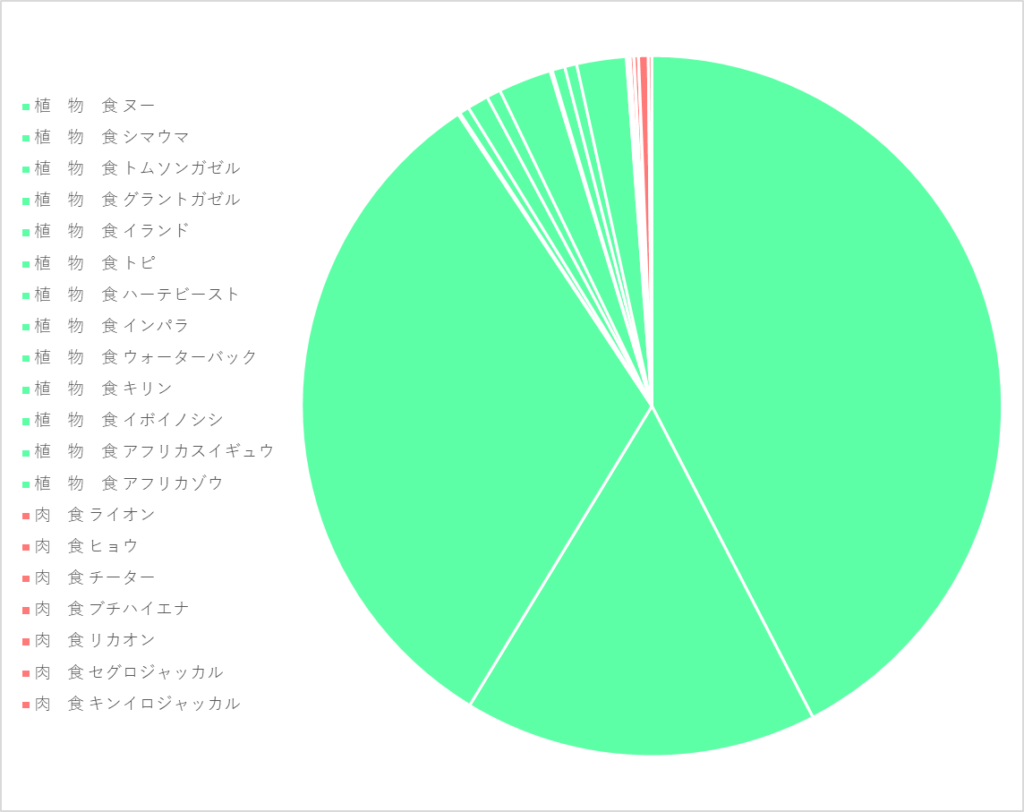
また、タンザニア*にあるセレンゲティ国立公園*という自然保護区*に生息する哺乳類の内、植物食の仲間はヌー*をはじめとして13種が確認され、数もヌーだけで約130万頭いるのに対し、肉食の仲間はライオン*をはじめとして7種、数に至っては全種の合計で約3.3万頭と少なく、多くの哺乳類が植物を食料源にしていることがわかります。

恐竜の場合では化石として発見されている種や個体数が、当時の生態系をどこまで反映しているか判断するのは難しいものの、北米のヘルクリーク累層*における8つの恐竜の仲間の群集を調べた研究では、最も多い仲間がケラトプシダエ類*の約61%であり、次いで多いのがハドロサウリダエ類*の約23%と、どちらも植物食恐竜であり、肉食恐竜で最も多い仲間でもティラノサウリダエ類*の約4%と少なく、今も昔も植物食の動物の方が、多くいたことがわかります。
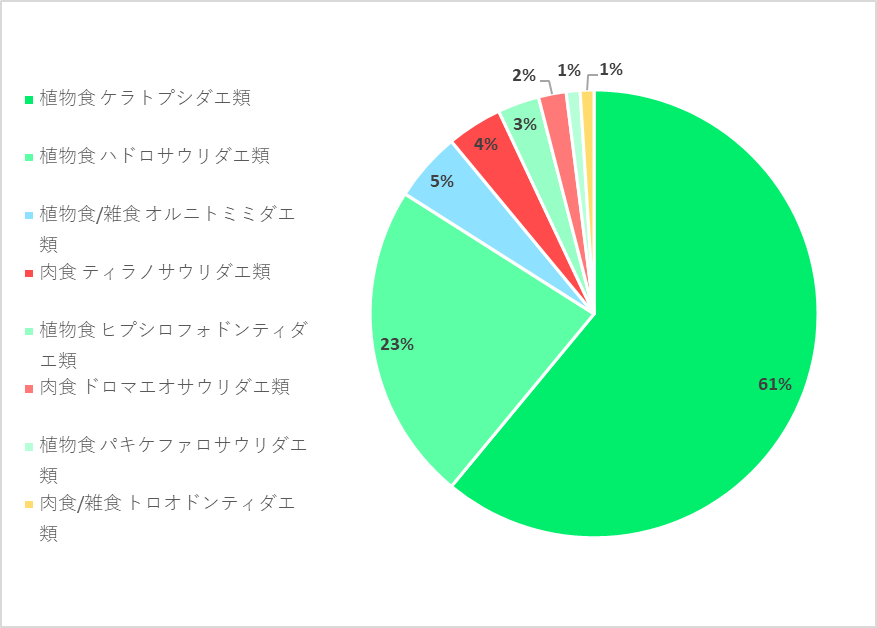
そもそも食物連鎖*の関係上、生産者*たる植物を食べる植物食動物よりも、その植物食動物を捕食する肉食動物の方が、数が少ないことは自然であり、食性という非常に大きな括りで比較しようとすると、多くの恐竜がウサギと共通するのは当然のようです。
5).中生代と新生代のサラダ
これまでの章で、ウサギの食性である植物食という括りでは、恐竜にも多くの仲間がおり、様々な部分でそれらが判断されていることは、知ることができたと思います。ただ、一重に植物食といっても、動物によって利用している種類や部分は異なっており、ウサギの仲間では草や木の葉、木の皮、根、果実、種子を食べることが分かっています。そこで、ここからは、より具体的にウサギと恐竜の食性を比べていきたいと思います。

・献立を考える前に
ウサギが食す植物と、恐竜が食していた植物を比べる際には気をつけるべきことがあり、そのひとつが、今と昔の植生*の違いです。現在、私達が見ることができ、ウサギたちが利用している植物は、昔から変わらず生えていた訳ではありません。

恐竜が生きていた時代の前半である後期三畳紀*から、後期ジュラ紀*頃まではシダやソテツ、ベネチテス類*、針葉樹*などの裸子植物が生い茂り、現在優勢な被子植物は後半のジュラ紀末から白亜紀初頭に登場して勢力を拡大し、白亜紀の中頃に主要な植物になったようで、過去の植物がつくる光景は、現在とは異なっていたようです。


この様な環境の違いは植物食動物の食性にも反映され、現生動物の多くが主要な食料源としている被子植物は、植物食恐竜が生きていた時代によっては存在せず、登場後においても恐竜にとっては新しい資源であり、主な食料源ではなかった可能性があるのです。
・献立1:草
ウサギの仲間がよく食べる一般的に“草”と呼ばれているイネ科植物*が登場したのは白亜紀初頭の頃で、当時は水辺の植物であり、勢力を拡大して草原*といった環境をつくるのは恐竜絶滅後の世界のことです。ただ、新参者であったイネ科植物も恐竜は利用していたようで、インドの後期白亜紀の地層から発見された、ティタノサウルス類のものと考えられている糞化石からは、イネ科の種子の残骸が発見されています。

また、中国の前期白亜紀の地層から発見されたエクイジュブス*という恐竜の歯列から、イネ科植物の表皮の微細な化石が発見されており、比較的新しい植物であったイネ科植物も恐竜は食料として利用していたようです。

・献立2:トクサ
イネ科植物以外では、トクサや他の植物の若い部分をウサギは好んで食べるそうですが、恐竜においてはディプロドクス類やステゴサウルス類*、ノドサウルス類、アンキロサウルス類*、ハドロサウルス類などの内容物や、これら植物食恐竜のものと考えられる糞化石からはトクサやシダ、ソテツ、針葉樹、顕花植物*が発見されており、現生種と化石種の違いなどに目を瞑れば、似たような植物も食べていたと考えることはできます。

・献立3:木
ウサギの仲間は草やトクサのほかに、主な食料源ではないものの木の皮や根、果実、種子等も食べることが知られていますが、恐竜においてもこれらの植物を食料といていた研究・発見した例は知られています。後期ジュラ紀から後期白亜紀の植物食恐竜の糞化石からは木片が発見されており、より具体的なものでは、マイアサウラ*のものと推測されている糞化石に多くの木片が含まれていたことなどから、偶然の採食はなく、明確に木も食料としていたと考えられているようです。

・献立4:果実・種子
ウサギの仲間をはじめ、多くの動物に利用される果実や種子は、アンキロサウルス類の内容物から、種や果実のようなものが発見されているほか、アルゼンチンの中期ジュラ紀*の地層から発見されたイザベリサウラ*という鳥盤類からは、ソテツの種子が見つかっています。また、ソテツやイチョウ*を食べた恐竜が現生動物でも見られる種子散布*の役割を担っていたのではないかという研究もあり、植物食恐竜も植物を幅広く食料源といて利用していたほか、植物側も分配域の拡大に植物食恐竜を利用していたのかもしれません。
・献立5:肉
最後に例外的ではありますが、肉を食べる場合について見ていきます。ウサギの仲間は、餌の不足で仲間の死骸や、出産後の栄養失調が原因で子供を食べる共食い*をするほか、鳥や天敵となる捕食者などの死骸を食べる事例が報告されているそうです。

恐竜においては、肉食恐竜で共食いの可能性が発見、研究されていますが、植物食恐竜内での共食いにいての事例は見つけることができませんでした。ただ、動物質の物を食べたという証拠はあり、ハドロサウルス類の糞化石から甲殻類*の破片や軟体動物(二枚貝)*の殻が見つかっています。これらは複数の糞化石から発見されている等の理由から、偶然ではなくタンパク質*や、カルシウム*の摂取が目的で意図的に食したと考えられているようです。

こういった不足した栄養素の補充のために、植物食動物が動物質の物を食べる行為は、ウサギ以外の現生動物にも見られ、有名な例では多摩動物公園のキリンがハト*を食した写真が、学研の図鑑「動物」に掲載されていたこともあります。また、野生下でもキリンがスイギュウ*の骨を食べる行為が確認されているほか、日本でもヤクシカ*が仲間の死骸を食べる例が報告されています。

哺乳類以外の植物食動物では、鳥類のマガモ*の仲間やオウム*の仲間において、植物食の傾向は強いものの、昆虫や軟体動物、魚類なども食べることが知られています。また、爬虫類においても植物食の傾向が強いリクガメ*の仲間が死骸を食べるほか、鳥の雛を捕食した例が報告されているようですし、イグアナの仲間においては、同種を捕食した例もあるようです。化石記録や現生動物の例を考えると、植物食恐竜たちも竜脚類や鳥盤類、植物食の獣脚類などそれぞれの仲間で、肉を食べる機会は以外と多かったのかもしれません。

6).最後に
これにてウサギと恐竜を比べるシリーズ、第2回「ウサギの食べ物と、恐竜との比べっこ」は終了となります。化石種と現生種の違いや、過去の環境や生態系など様々な条件を無視したほか、一種類の恐竜や系統に絞らずに、ウサギの食性と比較したりと、かなり強引な話の展開でしたが、如何だったでしょうか。
植物食という大きな括りだけではなく、細かい食性においいても似ている点はあったのではないか思います。今も昔も植物食の動物たちは、その時代、地域に自生する植物を資源として利用して生き、植物を食料とする限り、生物として遠い関係だったとしても、似た食性になるのは当然のことなのかもしれません。
【名前】
※前回紹介した名前に関しては省略しています。
・アジアゾウ:インドと東南アジアに生息するゾウ科の哺乳類で、さらにインドゾウとセイロンゾウ、スマトラゾウの3つの亜科に細かく分けられている。アフリカゾウと比べて小柄で、耳も小さい。
・アフリカゾウ:現生の陸上哺乳類で、最大の動物であり、アフリカ大陸に生息する。マルミミゾウとサバンナゾウの2種に分けられている。アジアゾウでは頭部の山なりになっていている部分が2つであるに対し、1つであるほか、鼻先の突起が上下にあり、アジアゾウの上のみにある鼻とは異なる。
・アンキロサウルス類:アンキロサウリダエ類のことで、ノドサウルス類(ノドサウリダエ類)と共に鎧竜(アンキロサウリア類)にまとめられる恐竜である。鳥盤類に含まれ、体の多くの部分が皮骨のプレートやとトゲに覆われており、尾の先にハンマーを持つ方の鎧竜である。
・イグアナ:グリーンイグアナの通称で、中央アメリカに生息する植物食の爬虫類である。
・イグアノドン:イグアノドン類の鳥脚類。ヨーロッパの前期白亜紀の地層から発見されている。メガロサウルスと並び、世界最初に科学的に研究された恐竜としても有名である。
・イザベリサウラ:ステゴサウリア類のとも考えられている鳥盤類の恐竜。アルゼンチンの前~中期ジュラ紀の地層から発見されている。二足歩行だった可能性もあるが、足まわりの化石は未発見である。
・イチョウ:イチョウ類の落葉性の高木で、裸子植物の仲間である。化石記録はペルム紀まで遡るが、生き残っているのは1種のみで、遺伝学的な研究と考古学的な記録から、中国原産と考えられている。
・ウシ:ウシ属の植物食の哺乳類で、野生の仲間はユーラシア大陸やアフリカ大陸に生息し,短い首や一対の角などが特徴である。一般的に想像されるホルスタインは、ボス・オーロックとい種が家畜化された生まれたものである。
・エクイジュブス:ハドロサウリダエ類の鳥脚類で、中国の前期白亜紀の地層から発見されている。上顎の尾側付近の歯から珪化した植物の表皮が発見されており、その形状からイネ科に共通する特徴があったことが分かっている。
・オウム:オウム科の鳥類の総称で、植物食の傾向が強い雑食である。オーストラリア、ニュージーランド、ニューギニアの周辺に生息し、近い仲間のインコとは冠羽を持つことや、緑や青色をみせる羽根の構造を持たないことで区別できる。
・オルニトミモサウルス類:全体的にはダチョウに似た体型の獣脚類の仲間であり、前肢の指の長さがほぼ同じなことや、よく発達した後肢の筋肉、大きい眼窩等の特徴を持つ。コエルロサウルス類という明確に羽毛と呼べる構造を持つようになった仲間に含まれ、ユーラシア大陸や北アメリカの白亜紀の地層で発見されているが、アフリカ大陸のジュラ紀の地層からも仲間と思われる化石が見つかっている。
・カメ類:カメ目に含まれる爬虫類の総称で、南極大陸を除く全ての大陸に生息するほか、インド羊、大西洋、太平洋などにも生息する。最も古い仲間は後期三畳紀に確認されており、発生学や分子系統学的な研究から、恐竜類やワニ類と同じ主竜類に含まれると考えられている。頭が甲羅にしまえる種類が出現するのは、前期白亜紀が最も古い化石記録である。
・キリン:キリン属の植物食哺乳類であり、長い首が特徴的であるが、多くの哺乳類と同じで頸椎の数は7個である。現生種は1種のみとされているが、亜種を独立した種とする考えもある。アフリカ大陸に生息するが、化石はユーラシア大陸からも発見されている。
・ケラトプシダエ類:ケラトプス科のことで、セントロサウリナエ類(セントロサウルス亜科)とカスモサウリナエ類(カスモサウルス亜科)をまとめた角竜の仲間のことで、大型化する外鼻孔や2つに分かれる歯根等で特徴付けられる。前者はフリルをつくる鱗状骨がハの字になり、外鼻孔の穴が1つである。後者は鱗状骨がL字になり、外鼻孔の穴が2つになる。
・コケ:コケ植物のことで蘚類、苔類、ツノゴケ類を含む非維管束植物の総称である。維管束を持たないことで特徴付けられ、胞子で繁殖するほか、無性生殖でも繁殖する。ユーラシア大陸、南北アメリカ大陸、アフリカ大陸、オーストラリア大陸、南極大陸と全ての大陸で見られ、化石では前期オルドビス紀のものが最古の記録として知られている。
・サイ:サイ科に含まれる植物食哺乳類の総称であり、現生種ではシロサイとクロサイがアフリカ大陸に、インドサイがインドとネパールに、スマトラサとジャワサイがマレーシアとインドネシアに生息する。頭が大きく、骨芯のない角質のツノが生えることや硬い皮膚等が特徴的である。化石では前期始新世のものが最古の記録として知られている。
・シダ植物:維管束植物の中で、胞子で繁殖する植物の総称であり、現生種は南極大陸を除く全ての大陸で見ることができる。化石では前期デボン紀のものが最古の記録として知られている。
・スイギュウ:アジアスイギュウとアフリカスイギュウは同じウシ亜科に含まれるものの、前者はアジアスイギュウ属、後者はアフリカスイギュウ属であり区別される。アフリカスイギュウは他の大型のウシ科と比べて胴長で、短く太い四肢をといった特徴を持っている。
・ステゴサウルス類:ステゴサウリア類のことで、皮骨板が棘状または板状に発達することや、四足歩行ながら後肢が前肢と比べて長いといった特徴がある。前上顎骨歯を持つことや、皮骨板が小さく数が多いことで特徴付けられるファンヤンゴサウリダエ類と、前上顎骨歯がないことや、体の横方向に生えるトゲが消失していることなどで特徴付けられるステゴサウリダエ類に分けられる。
・ソテツ:窒素固定細菌と共生するソテツ科の裸子植物のことで、日本の固有種である。ソテツ科の現生種は中央アメリカから南アメリカ、アフリカ、東南アジアなどの亜熱帯から熱帯地域に見られるが、オーストラリアなどの温帯地域でも見られる。化石では後期石炭紀のものが最古の記録と考えられている。
・ゾウ:ゾウ科に含まれる植物食哺乳類の総称であり、現生種ではアジアゾウがインドと東南アジアに、マルミミゾウとサバンナゾウがアフリカ大陸に生息し、筋肉質の長い鼻や大きな耳が特徴的である。化石では後期中新世のものが最古の記録として知られている。
・ティタノサウルス形類:外鼻孔が眼窩よりも大きいこと等で特徴付けられるマクロナリア類の中でもブラキオサウリダエ類とティタノサウリア類を含む竜脚類の仲間のこと。ブラキオサウリダエ類はマクロナリア類の中でも特に前肢が長いことや、中手骨が細長いことで特徴付けられ、ティタノサウリア類は橈骨と尺骨が太いことや、頭が他の竜脚類より小さいといった特徴を持つ。
・ティタノサウルス類:ティタノサウリア類のことで、エウへロプス類と共にソムフォスポディリ類をなすティタノサウルス形類の竜脚類である。腸骨が前方に向かって広がるといった特徴を持ち、南北アメリカ大陸、ユーラシア大陸、アフリカ大陸、オーストラリア大陸、南極大陸と全ての大陸で化石が発見されている。
・ティラノサウリダエ類:アルバートサウリナエ類とティラノサウリナエ類からなる獣脚類の仲間であり、北アメリカとアジアの後期白亜紀の地層から発見されている。方形頬骨と鱗状骨の縫合部が、外側頭窓の中に強く張り出すとった特徴を持ち、ティラノサウリナエ類には有名なティラノサウルスも含まれる。
・トクサ:トクサ科の植物のことであり、北半球の亜寒帯から温帯に広く分布する。同じ仲間のスギナ等とは異なり、枝分かれせずに中空の節をつくる。化石では前期石炭紀のものが最古の記録として知られている。
・ヌー:ウシ科に含まれる植物食哺乳類であり、ケニアやタンザニアなどアフリカ大陸の中央部や南部、東部に生息し、オグロヌーとオジロヌーが存在する。ウシとカモシカを合わせたような形態をしていることや、大きな群れをつくることで知られている。
・ハト:ハト科に含まれる雑食性の鳥類の総称であり、一部の砂漠や極域と周辺の島々を除くあらゆる地域に分布する。日本ではキジバトが最も数が多く一般的であり、次いでドバトが良く知られているが、その他にもシラコバトやアオバト、カラスバトなどがいる。
・ハドロサウリダエ類:ハドロサウルス科のことで、サウロロヒナエ類(サウロロフス亜科)とランベオサウリナエ類(ランベオサウルス亜科)をまとめたハドロサウルス類の仲間のことで、平たく横に伸びた口先や発達したデンタルバッテリーで特徴付けられる。前者は骨質で中空のトサカを持たず、後者はトサカがよく発達する。
・バクテリア:細菌のことであり、外部からエネルギーを取り入れて毒素や水分を排出する単細胞の微生物で、酸素をつくることで有名なシアノバクテリアは化石としては約37億年前のものがグリーンランドから発見されている。様々な環境に適応しており、他の生物に寄生するものも存在するが、全てが有害という訳ではなく、中には有益なものも存在する。
・パキケファロサウリア類:堅頭竜類のことで、厚く発達した前頭骨と頭頂骨で特徴付けられるが、必ずしも有名なパキケファロサウルスの様にドーム状になる訳ではなく、平たく発達するホマロケファレといった仲間も存在する。
・パキケファロサウルス類:パキケファロサウリダエ類(パキケファロサウルス科)のことで、堅頭竜類の下部に分類されるが、現状では中国で発見されている原始的な仲間を除いて、全てがパキケファロサウルス類に含まれているようである。
・フクイサウル:イグアノドン類の中でもアンキロポレックス類に含まれる鳥脚類の恐竜であり、福井県の前期白亜紀の地層から発見させている。一般的なイグアノドン類とは異なり、上顎のキネシスを失っていたほか、下顎が分厚くなっているのが特徴である。因みに、日本で最初に国産の復元骨格がつくられた恐竜である。
・フクイティタン:ティタノサウルス形類に含まれる竜脚類の恐竜であり、福井県の前期白亜紀の地層、手取層群北谷層から発見されている。前肢に指骨がないタイプの竜脚類であると、足跡の研究から考えられている。因みに、日本で最初に学名が付けられた竜脚類である。
・フクイラプトル:アロサウルス上科の中でもメガラプトル類に含まれる獣脚類の恐竜であり、上記の2つの福井県産の恐竜と同様の発掘現場である、手取層群北谷層の赤岩亜層群から発見されている。メガラプトル類の中では原始的であるものの、この仲間に見られる大きく薄い末節骨が特徴的である。因みに、日本で最初に学名が付けたれた恐竜である。
・ベネチテス類:裸子植物の一種であり、後期三畳紀に登場して白亜紀末に絶滅した中生代の植物で、化石は世界中から見つかる。見かけはソテツに似ているとされているが、近年の日本を含む保存状態の良い化石から従来考えられていたほどソテツに似た見た目ではにない可能性がある。
・ボレアロペルタ:ノドサウルス類に含まれるヨロイ竜の恐竜であり、カナダの前期白亜紀の地層から発見されている。軟組織に関連する保存が良い所謂ミイラ化石であり、皮骨の装甲の位置がほぼ生前をとどめていたほか、皮骨を覆うケラチン質やウロコからはメラノソームも発見されている。
・マイアサウラ:サウロロフス亜科に含まれるハドロサウルス類の恐竜であり、アメリカの後期白亜紀の地層から発見されている。成体と幼体を含む巣が近い所で発見されたことや、当時の研究で幼体の足が歩行に十分な形態になっていなかった事などから、初めて子育ての可能性が指摘された恐竜である。ただし、幼体には十分な歩行能力があったという研究もあり、子育てについては慎重に判断されるべきである。
・マガモ:カモ科に含まれる雑食性の鳥類であり、北半球の亜寒帯から温帯に広く分布し、日本では秋に北から渡ってきて、春に北へ戻る冬鳥として全国的に見ることができる。北海道等の一部の地域では繫殖も行う。
・ヤクシカ:ニホンジカの亜種に含まれる基本的には植物食の哺乳類であり、鹿児島の屋久島と口永良部島のみに生息している。ニホンジカと比較して小型であることや、オスの角の枝分かれが4本から3本に減っていることなどで特殊付けられる。
・ライオン:ヒョウ族に含まれるネコ科の肉食動物であり、現生種の中ではトラに次ぐ大型のネコ科で、アフリカ大陸およびインドに分布する。群れをつくる珍しいネコ科であり、定住性のプライドと呼ばれメスを中心とした繫殖も行う群れが有名である。また、コアリションと呼ばれるオスの連帯も存在する。
・リクガメ:リクガメ科に分類されるカメの総称であり、ユーラシア大陸やアフリカ大陸、南北アフリカ大陸など分布する植物食の傾向が強い雑食性のカメである。現生種には甲長130㎝を超えるものから10㎝に満たない種まで存在する。
【用語】
※前回紹介した用語に関しては省略しています。
・イネ科植物:被子植物の一種で、南極大陸などの全大陸、乾燥地帯から高山帯まで世界中のあらゆる環境に広く分布しており、8000種以上の種がいるなど非常に成功している仲間である。一つの花に一つの種子をもちつつ、花の数を増やしているため、十分な栄養を持った種が沢山つくられる優れた繫殖戦略を取る。
・カルシウム:熱に強く水に溶けにくいアルカリ土類金属に分けられる金属で、生物の体を作り、筋肉や神経の反応に関わる身体にとって重要な要素のことである。自身でミネラルをつくることができないため、食物から摂取する必要がある。カルシウムは骨や歯の形成に用いられるほか、出血時には血液の凝固を促進する効果がるなど、生物に最も多く含まれるミネラルである。
・セレンゲティ国立公園:アフリカ大陸のタンザニア北部に存在する生物圏保護区に登録している国立公園のことで、多数の動物が生息しており、自然の美しさと生物の多様性から世界自然遺産に1981年から登録している。ライオンやヌー等の哺乳類からダチョウやコシジロハゲワシ等の鳥類、ナイルワニやニジトカゲなどの爬虫類など様々な動物を見ることができる。因みに、セレンゲティとはマサイ族の言葉で「果てしなく広がる草原」を意味する。
・タンザニア:タンザニア連邦共和国のことで、ドドマを首都とするアフリカ大陸の東に位置する国である。セレンゲティ国立公園などの自然が知られる一方で、100を超える原住民族とアラブ文化とヨーロッパ文化が混ざる文化的にも興味深い国である。恐竜化石では中期ジュラ紀から前期白亜紀の地層が広がるテンダグル層の恐竜たちが有名であり、ケントロサウルスやジラファティタンなどが知られている。また、中期三畳紀のマンダ層からは最古の恐竜類に含まれる可能性のあるニャササウルスも発見されている。
・タンパク質:アミノ酸の高分子化合物のことで、筋肉や臓器など生物体のあらゆる組織をつくる材料となるほか、ホルモンや酵素、神経伝達物質の材料ともなる。タンパク質は基本的には生物固有の物質であり、合成が細胞の中で行われる。しかし、生命誕生前の地球ではタンパク質が非生命的に合成された後、核酸と相互作用して生命に化学的に進化したと考えられている。
・ヘルクリーク累層:北アメリカにある後期白亜紀から新生代暁新世の地層のことで、アメリカのモンタナ州やノースダコタ州、サウスダコタ州、ワイオミング州に存在する。白亜紀末期の恐竜たちの化石が発見されていることで有名であり、ティラノサウルスやトリケラトプス、アンキロサウルス、パキケファロサウルスなどが知られている。
・臼歯:哺乳類の犬歯より後ろの歯のことであり、小臼歯と大臼歯に分けることができ、小臼歯は食物を噛み砕く役割を、大臼歯はすり潰す役割を持つとされている。肉食動物の場合には臼歯も尖った形状をしており、肉を裂き、骨を噛み砕くのに用いられ、植物動物の場合には平たく葉をすり潰すのに用いられる。雑食性の場合には両者の中間的な形状を示す。
・栄養失調:体に必要なエネルギーやタンパク質、ビタミンなどを充分に補給できていない状態のことであり、食事の量的な問題から偏食、病気等によって引き起こされる。動物では環境変動に起因するホッキョクグマの栄養失調が有名である。
・学名:世界共通で付けられる生物の分類群の名称のことで、国際命名規約という国際的なルールに則って付けられる。動物の場合には国際動物命名規約に従い、新種と思しき動物を発見した場合には、似ている種の論文と比較して一致しない時に新種の記載論文を書き、公表することで新種として認められる。動物の場合には属名と種小名を付け、新種または新属新種として公表れ、恐竜のような古生物も同じルールに乗っ取り名付けられる。古生物の場合には現生動物に用いられる幾つかの研究手法が使えない場合もあるため、新属新種になることが多い傾向にある。
・共食い:同種の動物間で一方が他方をたべることであり、濾過摂食をする魚が偶然に幼体や卵食べてしまう偶発的なものや、カマキリのように交尾の後にメスがオスを食べるような繫殖に伴うもの、ライオン等で見られる親の栄養状態が悪い場合に死んだ子供や、生き残る可能性の低い子供を食べるもの、クモで見られる母体が子供の餌となるもの等、様々なものが存在する。
・剣竜:ステゴサウリア類と呼ばれ、皮骨でできたプレートやスパイクが背中から尻尾にかけて発達する鳥盤類の恐竜であり、体の多くの部分がプレートやトゲに覆われていたアンキロサウリア類(鎧竜)と、他の原始的な仲間を合わせて装盾類というグループにまとめられる。剣竜はファンヤンゴサウリダエ類とステゴサウリダエ類に大きく分けられる。
・顕花植物:花咲かせて種子をつくる有性生殖の植物のことで、種子植物のことである裸子植物と被子植物の両方が含まれ、植物の多くがこれに該当する。最古の化石記録は約3億8500万年前の中期デボン紀のベルギーの地層から発見されているルンカリアだとされている。
・後期ジュラ紀:地球の地質時代のひとつである中生代を三つに分けた時に真ん中の時代がジュラ紀であり、更にジュラ紀を三つに分けた内、最も新しい時代のことである。約1億6150万年前から約1億4500万年前のこと。
・後期三畳紀:地球の地質時代のひとつである中生代を三つに分けた時に最初の時代が三畳紀であり、更に三畳紀を三つに分けた内、最も新しい時代のことである。約2億3700万年前から約2億140万年前のこと。
・甲殻類:体をキチン質の固い殻で覆い、節のある肢を持つ節足動物のうち、エビやカニ、ヤドカリ等の内肢と外肢に二股に分かれた肢をもつ仲間のことである。100万種以上の仲間がいる節足動物の中で約67000種が甲殻類に含まれ、淡水と海水を中心に生息している。最古の化石記録は約5億年前のカンブリア紀のカナダやロシア、中国、ポーランドの地層から発見されているオルステン動物群の中から発見されている節足動物だとされている。
・行動観察:動物の行動を機能や機構、発達と進化といった観点から、研究対象の中で知りたいことを定め、動物の行動を記録しいてデータの収集と解析を行う研究の方法のことである。客観的で再現性のあるデータを収集することが重要であり、行動を予め定義付けて、それに従って一定の規則の下でデータを収集する。
・採食:動物が自身で食べ物を摂取することであり、肉食や植物食に問わず用いられる。同様の意味をもつ言葉には「採餌」や「摂食」などがある。
・細胞壁:細胞膜の外側を囲っている細胞外基質のことであり、植物に見られるほか、キノコなどの真菌類やシアノバクテリアなどの細菌類でも見ることができる。
・歯冠:歯のなかでも口内に露出している部分のこと。
・歯根:歯茎などに埋まっていて見えない歯冠より下の部分のこと。
・歯列:歯の種類ごとの配列のこと。
・自然保護区:自然を守るために人間活動を控える地域のことで、様々な種類が存在し、世界的に有名なものではタンザニア北部にあるンゴロンゴロ自然保護区が、日本では尾瀬国立公園などがある。
・種子散布:種子が母体から離れて移動することで、種子植物が生育環境を拡大する際には注意行われる。どんな形に種子が運ばれるかといった単位や、どの様に運ばれるといった方法には様々な種類があり、種子や果実の形態に関係している。
・植生:植物の集団の分布を示した地図のことで、現存植生や潜在自然植生などが存在する.また優占種等に着目して相観植生や、植物群落の組成に着目して区別する方法などがあります。
・植物食:植物を主な食物として生きる性質のこと。
・食物連鎖:生物で行われる食う・食われるの関係のことで、二酸化炭素を固定して有機物をつくる生産者から始まり、それを食べる一次消費者、更にそれを食べる二次消費者と繋がる。最も上位の消費者は捕食者であり、これに至るまでの長さを食物連鎖長という。また捕食者も死後はバクテリアなどに分解されていく。
・針葉樹:葉が針の様に細長く堅い、マツやスギなどの裸子植物球果植物のことを一般的に示すが、被針形など針様ではない葉をつける種類もいる。熱帯から温帯、ツンドラのような寒帯まで広く分布している。広葉樹との見極めかたとしては幹を見るのが有効であり、針葉樹は真っ直ぐ伸びるのに対し、広葉樹は曲がったり,分化しているほか,内部の構造も異なっている。
・生産者:無機物から有機物をつくる生物のことで、太陽の光を受けデンプン等をつくる植物や海藻、植物プランクトン等が該当する。食物連鎖におけるエネルギーを作り出している側であり、これを食べる植物食動物や,植物食動物をたべる肉食動物等の消費者は、生産者の作る栄養を生物間で受け渡して生命を維持している。
・前歯骨:鳥盤類の恐竜に共通する特徴であり,下顎の先端,歯骨の前にある骨のことで,歯は生えておらず,クチバシが発達していた。
・草原: 草本植物を主とする群落のことで、ススキのようなイネ科の草が多く、地表面の半分以上が植物で覆われている場合や、低木が少数を混じる場合もある。森林の成立ができない様な高湿や乾燥、低温の地域に発達し、構成する植物の種類相や立地によって、イネ科草原、湿地草原、高層湿原等に分けられる。
・第1胃:ルーメンとも言われるウシ等の反芻動物がもつ4つの胃の内、一番目の胃のことで、焼き肉等ではミノにあたる部分のことである。4つの胃の中で最も大きく、成牛では胃全体の80%を占めており、容積は約200Lと大きい。この中には多数の微生物が共生しており、その微生物の作用によって植物の繊維を栄養として吸収できる形に分解している。
・恥骨:骨盤を構成する骨盤の骨の一つで、内臓を守り、体重を支える役割を持ち、爬虫類と哺乳類では寛骨と仙椎とは癒合しないことが多く、鳥類では成長とともに癒合して一体化して腹側で大きく開いた1個の骨盤となる。恐竜では多くの爬虫類の骨盤と同じ様に形恥骨が前下方を向いて者が多い仲間を竜盤類、恥骨が後ろに伸び、座骨と平行になっている仲間を鳥盤類として分けている。
・中期ジュラ紀::地球の地質時代のひとつである中生代を三つに分けた時に真ん中の時代がジュラ紀であり、更にジュラ紀を三つに分けた内、真ん中の時代のことである。約1億7470万年前から約1億6150万年前のこと。
・腸骨:骨盤を構成する骨盤の骨の一つで、骨盤の前背部を占める部分で脊柱に連結する部分のこと。 腿を動かし、骨盤の位置を正常に保つ働きを持つ骨盤と脚の付け根を結ぶ筋肉等が付着する。
・内部共生生物:多くの生物が恒常的もしくは、半恒常的に他の生物を体内にすまわせており、これを内部共生という。内部共生はほとんどの場合は微生物であり、空間的な近接性で成立する共生関係のため、高度な相互作用や依存関係がみられる。
・軟体動物:背骨を持たない無脊椎動物と言われる動物の中の一つで、同じ無脊椎動物の中では節足動物に次いで大きいグループで10万種以上いると推定されている。骨も節もないため文字通り柔らかな体をしており、外套膜という体の表面を覆う役割を持つ器官を持つという特徴があり、イカやタコの胴体部分や、貝類の内臓を包んでいる薄い膜がそれにあたる。
・二枚貝:石灰質の外骨格、いわゆる貝殻を左右二枚にもつ軟体動物のことで、体は頭部や筋肉質の発達した足、内臓塊でできており、口内には歯舌というおろし金状の摂餌器官をもつという特徴がある。足が斧のような形をしていることから斧足類とも言われ、海水や淡水、汽水域など生息し、二万種以上いるとされる。
・発酵器:一般的にはパンの生地を発酵させるためのもので、温度や湿度を一定に保ってパン生地を適正な状態で発酵させることができる。生物では、植物食動物が消化し辛い硬い繊維質を持つ植物を消化するために、微生物の力を借りて繊維を発酵させ消化し、栄養を吸収しており、消化管内に微生物を棲まわせた発酵槽をもっている。
・反芻動物:植物動物の中でも1度飲み込んで胃で消化した食べ物を口に戻し、繰り返し咀嚼する反芻という消化方法を取る動物のこと。四つの胃をもつことが特徴であり、ウシやヤギ等が有名である。
・被子植物:胚珠という種子の元になる部分が、子房という果実の元になる物に覆われることで守られている植物のこと。花をつけて受粉後に果実をつくり、果実は種子を包むことで保護するほか、動物に食されることで種子を遠くに運ぶ役割を果たす。
・被捕食者:生物で行われる食う・食われるの関係の内、食べられる側の生物のこと。生態系の中では、食う側である捕食者は被捕食者であることも多い。
・糞化石:動物の排泄物が化石化したもので糞石とも言い、地層中に残った生き物の行動の痕跡であることから、生痕化石のひとつとされる。形から糞の落とし主の排せつ口の形がわかるほか、未消化の食べ物が残されている場合、何を食べていたかある程度推定できる。
・糞分析:採取した糞を洗い、残された内容物の頻度を調べることで、その動物の食性を調べる方法のこと。虫の脚が1つの糞から大量に出たりすると数えるのが辛い。
・捕食者:生物で行われる食う・食われるの関係の内、食べる側の動物のこと。被捕食者の量が増えると植物が減っていき、生態系のバランスが崩れることから、頂点捕食者は被捕食者の数を調整する役割を担っているとされる。
・盲腸便:柔らかくて粘液に包まれたブドウの房状の糞で、栄養価の低い植物を食べるウサギは、盲腸内にいる微生物によって作られた便を食べることで、必要な栄養素を得ている。盲腸便にはビタミンやタンパク質が含まれている。
・木炭:一般的には木材を蒸し焼きにして得られる炭のことで、酸素が少ない状態でつくられる。森林火災後に樹木が育つには、火事発生時に作られる炭が重要な役割を果たしているとされるほか、微生物やミミズなどの土壌動物の活動を活性化し、土壌の養分循環へも大きく影響しているとされる。
・裸子植物:種を作る植物の内、胚珠が裸出している植物のこと。胚珠や花粉は葉の表面に形成され、胚珠が形成される葉は専用に変化してや花へと発展する。
・竜盤類:恐竜類を構成する二つの大きなグループのうちのひとつで、恥骨が前を向く骨盤が、現在のトカゲの骨盤に形が似ていたため名付けられた。肉食や植物、雑食など幅広い食性が見られ、肉食が多く鳥類も含まれる獣脚類と、原始的な仲間を除き植物食で、陸上最大級で長い首が特徴的な竜脚類の2つのグループに大きく分けられる。
・咀嚼筋:咀嚼に関係する筋肉の総称で、側頭筋や咬筋、外側翼突筋、内側翼突筋等がある。
・齧歯類:物をかじるのに適した歯と顎を持ち、上下顎の両方に伸び続ける2つの門歯があることや、犬歯を持たないことが特徴である。日本ではネズミ目とも呼ばれ、地球上で最も繁栄している哺乳類であり、南極大陸を除く全ての大陸に生息する。
【参考】
〈ネット〉
・NATIONAL GEOGRAPHIC 日本版. 「動物大図鑑 アジアゾウ」.2014.
https://natgeo.nikkeibp.co.jp/nng/article/20141218/428790/ (参照2023-01-06)
・NATIONAL GEOGRAPHIC 日本版. 「動物大図鑑 アフリカゾウ」.2014.
https://natgeo.nikkeibp.co.jp/nng/article/20141218/428791/ (参照2023-01-06)
・NATIONAL GEOGRAPHIC 日本版.「カメラは見た! 実は肉食系でもあるノウサギ」.2019.
https://natgeo.nikkeibp.co.jp/atcl/news/19/011600036/ (参照2023-01-05)
・Rob The Ranger Wildlife Videos. 「Rare Footage Of “Scavenging” Giraffe Herd Feeding On A Buffalo Skeleton! | Amazing African Wildlife」.2017.
https://youtu.be/pCy7dY8oREs (参照2023-01-06)
・WWFジャパン.「アジアゾウにつて」.2020.
https://www.wwf.or.jp/activities/basicinfo/4306.html (参照2023-01-06)
・WWFジャパン. 「地上最大の野生動物 ゾウ」.2020.
https://www.wwf.or.jp/activities/basicinfo/4291.html (参照2023-0106)
・屋久島自然研究会. 「シカが喰っていたものは・・・(自然の掟に弱い方は見ないで)」.2007.
http://canyon.air-nifty.com/forest/2007/12/post_097b.html (参照2023-01-06)
・研究.net. 研究用語辞典. 「食物連鎖」.
http://www.kenq.net/dic/168.html (参照2023-01-06)
・公益財団法人 東京動物園協会. 東京ズーネットBB. うごく!どうぶつ図鑑. 「アジアゾウ」
https://www.tokyo-zoo.net/encyclopedia/species_detail?code=2 (参照2023-01-06)
・公益財団法人 東京動物園協会. 東京ズーネットBB. うごく!どうぶつ図鑑. 「アフリカゾウ豆知識」.
https://www.tokyo-zoo.net/topics/profile/profile23.shtml (参照2023-01-06)
・公益財団法人 東京動物園協会. 東京ズーネットBB. うごく!どうぶつ図鑑. 「キリンの反芻」.2016.
https://www.tokyo-zoo.net/movie/mov_book/1603_02/index.html (参照2023-01-06)
・社会実情データ図鑑. 「アフリカ・サバンナの動物たち」.2009年7月1日収録.2020年10月14日図改良.
https://honkawa2.sakura.ne.jp/4176.html (参照2023-01-06)
・独立行政法人 農畜産業振興機構. 広報誌. 「【まめ知識】なぜ、牛の胃は4つもあるの?」
https://www.alic.go.jp/koho/kikaku03_001316.html (参照2023-01-06)
〈書籍〉
・D. E. Fastovsky・D. B. Weishampel.『恐竜学入門』.東京化学同人,2015,400p
・Darren Naish・Paul Barrett.『恐竜の教科書』.創元社,2019,240p
・David Norman.『恐竜 化石記録が示す事実と謎』.丸善出版株式会社,2014,222p
・Edwin Harris Colbert.『恐竜の発見』.早川書房,1993,444p
・G.Masukawa.『新恐竜骨格図集』.イースト・プレス,2022,160p
・Robert T. Bakker.『恐竜異説』.平凡社,1989,326p
・加藤嘉太郎. 山内昭二.『新編 家畜比較解剖図説 上巻』.養賢堂,2007,315p
・吉田賢治.『頭骨大図鑑Skulls』.アプリスタイル,2021,256p
・恐竜戦国時代の覇者!トリケラトプス~知られざる大陸ララミディアでの攻防~.大阪市立自然史博物館,2014.
・恐竜博2019.国立科学博物館,2019年.
・犬塚則久.『恐竜ホネホネ学』.日本放送出版協会,2006,261p
・獣脚類 ―鳥に進化した肉食恐竜たち―.福井県立恐竜博物館,2018年.
・土屋健.『生物ミステリー こっそり楽しむうんこ化石の世界』.技術評論社,2022,239p
・富田京一.『ホネホネ動物ふしぎ大図鑑』.日本図書センター,2018,164p
〈論文〉
・Attila Ősi, Paul M. Barrett., Alistair R. Evans., András Lajos Nagy., Imre Szenti., Ákos Kukovecz., János Magyar., Martin Segesdi., Kinga Gere., Viviána Jó., 2022. Multi-proxy dentition analyses reveal niche partitioning between sympatric herbivorous dinosaurs. Scientific Reports, volume 12, Article number 20813.
・Caleb M. Brown., David R. Greenwood., Jessica E. Kalyniuk., Dennis R. Braman., Donald M. Henderson., Cathy L. Greenwood., James F. Basinger., 2020. Dietary palaeoecology of an Early Cretaceous armoured dinosaur (Ornithischia; Nodosauridae) based on floral analysis of stomach contents. Royal Society Open Science, 7(6), 200305.
・DAVID J. BUTTON., PAUL M. BARRETT., EMILY J. RAYFIELD., 2016. COMPARATIVE CRANIAL MYOLOGY AND BIOMECHANICS OF PLATEOSAURUS AND CAMARASAURUS AND EVOLUTION OF THE SAUROPOD FEEDING APPARATUS. Palaeontology, Vol. 59, Part 6, p887–913.
・GREGORYM.ERICKSON.,BRANDONA. KRICK., MATTHEW HAMILTON., GERALD R. BOURNE., AND W. GREGORY SAWYER., 2012. Complex Dental Structure and Wear Biomechanics in Hadrosaurid Dinosaurs. SCIENCE, Vol 338, Issue 6103, p98-101.
・George L. W. Perry., 2021. How for might plant-eating dinosaurs have moved seeds?. THE ROYAL SOCIETY, Biol. Lett. 17: 20200689.
・Homare Sakaki., Daniela E. Winkler., Tai Kubo., Ren Hirayama., Hikaru Uno., Shinya Miyata., Hideki Endo., Kazuhisa Sasaki., Toshio Takisawa., Mugino O. Kubo., 2022. Non-occlusal dental microwear texture analysis of a titanosauriform sauropod dinosaur from the Upper Cretaceous (Turonian) Tamagawa Formation, northeastern Japan. Cretaceous Research, Volume 136, 105218.
・Justin S. Tweet., K. Chin., Nate Murphy., 2008. Probable Gut Contents Within A Specimen Of Brachylohosaurus Canadensis (Dinosauria : Hadrosauridae) From the Upper Cretaceous Judith River Formation Of Montana. SEMANTIC SCHOLAR, p07-044.
・Karen Chin., 2007. THE PALEOBIOLOGICAL IMPLICATIONS OF HERBIVOROUS DINOSAUR COPROLITES FROM TWO MEDICINE FORMATION OF MONTANA: WHY EAT WOOD?. PALAIOS, Vol.22, No.5, p554-566.
・Karen Chin., Rodney M. Feldmann., Jessica N. Tashman., 2017. Consumption of crustaceans by megaherbivorous dinosaurs: dietary flexibility and dinosaur life history strategies. Scientific Reports, volume 7, Article number 11163.
・Leonardo Salgado., José I. Canudo., Alberto C. Garrido., Miguel Moreno-Azanza., Leandro C. A. Martínez., Rodolfo A. Coria., José M. Gasca., 2017. A new primitive Neornithischian dinosaur from the Jurassic of Patagonia with gut contents. Scientific Reports, volume 7, Article number 42778.
・Lindsay E. Zanno., Peter J. Makovicky., 2010. Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. PNAS, 108(1), 232-237.
・Paul D. White., David E. Fastovsky., Peter M. Sheehan., 1998. Taphonomy and Suggested Structure of the Dinosaurian Assemblage of the Hell Creek Formation (Maastrichtian), Eastern Montana and Western North Dakota. PALAIOS, Vol. 13, No.1, p41-51.
・Steven R Manchester., Thomas M. Lehman., Elisabeth Wheeler., 2010. Fossil Palms (Arecaceae, Coryphoideae) Associated with Juvenile Herbivorous Dinosaurs in the Upper Cretaceous Aguja Formation, Big Bend National Park, Texas. International Journal of Plant Sciences, 171(6), 679-689.
・Vincent S. Williams., Paul M. Barrett., Mark A. Purnell., 2009. Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding. PNAS, 106 (27), 11194-11199.
・Yan Wu., Hai-Lu You., Xiao-Qiang Li1., 2018. Dinosaur-associated Poaceae epidermis and phytoliths from the Early Cretaceous of China. National Science Review, Volume5, 721–727.
・Yoshitsugu Kobayashi., Jun-Chang Lu., Zhi-Ming Dong., Rinchen Barsbold., Yoichi Azuma., Yukimitsu Tomida., 1999. Herbivorous diet in an ornithomimid dinosaur. Nature, volume 402, pages 480–481.
・Z. Dt. Ges. Geowiss. (J. Appl. Reg. Geol.)., 2020. A reappraisal of the “stomach” contents of the Edmontosaurus annectens mummy at the Senckenberg Naturmuseum in Frankfurt/Main (Germany). Zeitschrift der Deutschen Gesellschaft für Geowissenschaften Band 171 Heft 1, p71–85.
〈その他〉
・宇部尚樹. 石原亨.「イネ科植物における二次代謝の進化」.日本農芸化学会. 2018, 化学と生物, Vol.56, No.2, p. 73-75
・福江佑子. 竹下毅. 中西希. 「食肉目の研究に関わる調査技術事例集」食肉目における食性研究とその方法 その1 ―イヌ科,イタチ科,ネコ科―. 2011, 日本哺乳類学会. 哺乳類科学51(1):129-142.
【登場した博物館関連施設】
・Royal Tyrrell Museum of Palaeontology
・東海大学自然史博物館(閉館)
次回予告
第3回:うさぎの行動・生態と、恐竜との比べっこ ~ 恐竜死して兎悲しむ? ~